Lifespan Age Differences in Memory Representations (LIME)
Research Scientists
Myriam C. Sander (on maternity leave 07/2021–03/2023)
Anna E. Karlsson (until 02/2022)
Ulman Lindenberger
Claire Pauley
Verena Sommer (until 09/2021)
Research Area 1: Age Differences in Similarity and Distinctiveness of Memory Representations
Research Area 2: Effects of Context on Memory Representations
LIME addresses mechanisms of memory formation, consolidation, and retrieval, with a focus on the ways in which these mechanisms change across the lifespan (see Sander et al., 2021). In particular, the project investigates whether aging-induced decrements in the distinctiveness of neural representations contribute to age-related losses in memory performance in later adulthood. Providing answers to this question requires the coordination of concepts and methods from lifespan psychology, cognitive neuroscience, and computational neuroscience. Accordingly, the project relies on experimental research designs, advanced multi-modal imaging methods, and computational modeling. The project has been partially funded as a Minerva Research Group awarded to Myriam C. Sander.
Research Area 1: Age Differences in Similarity and Distinctiveness of Memory Representations
Are memories represented differently in older adults compared to younger adults? According to the dedifferentiation hypothesis, age-related cognitive decline during adulthood and old age reflects decrements in the distinctiveness of neural representations and processing pathways (Li et al., 2001). In line with this hypothesis, pioneering work by Denise Park and colleagues has shown that differences in the neural representation between items belonging to different categories, such as houses and faces, are less pronounced in older adults than in younger adults. However, the relation between differences in distinctiveness of memory representations and adult age differences in cognitive performance continue to be poorly understood. Furthermore, most research has investigated age differences in the distinctiveness of representations during memory formation, but rarely have age differences in distinctiveness been studied during memory retrieval. In LIME, we examine whether links between neural distinctiveness and performance help to elucidate adult age differences in episodic memory by following memory representations from encoding to retrieval (Sander et al., 2021).
For instance, in one of our recent studies we made use of functional magnetic resonance imaging (fMRI) during both the encoding and retrieval phases of a memory task (Dissertation Claire Pauley). Using exploratory representational similarity searchlight analyses, we identified brain regions showing an age-related deficit in neural distinctiveness during encoding, retrieval, and in encoding-retrieval reinstatement (see Figure 1). Specifically, we found that neural representations in occipital and ventral visual regions were less distinct in older adults compared to younger adults, and that lower distinctiveness across all phases was associated with poorer memory performance. Interestingly, interindividual differences in retrieval- and reinstatement-related neural distinctiveness were closely tied to distinctiveness during encoding. Furthermore, neural distinctiveness during encoding predicted individual differences in memory performance to a far greater extent than either retrieval- or reinstatement-related distinctiveness, in line with the proposition that age-related differences in encoding show downstream effects on later processing stages (Sander et al., 2021). In ongoing analyses, the project aims to understand how age differences in the connectivity of large-scale brain networks contribute to neural dedifferentiation (in cooperation with Dasa Zeithamova, visiting researcher in 2021).
 than older adults during encoding (a), retrieval (b), and reinstatement (c) for house stimuli.")
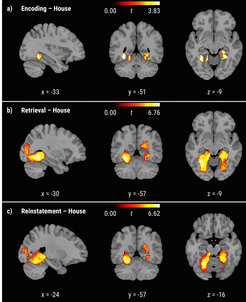
Image: MPI for Human Development
Adapted from Pauley et al. (2023)
Original image licensed under CC BY 4.0
In an EEG study, we investigated the detailedness of representations from childhood to old age (Dissertation Verena Sommer). Here, we used neural adaptation as a marker of the specificity of representations. This approach rests on the assumption that neuronal populations reduce their responses (i.e., adapt) when stimulus features to which they are sensitive are repeated. The magnitude of adaptation thus serves as an indicator of representation specificity. We asked whether differences in adaptation magnitudes predict differences in memory performance in children, young adults, and older adults. To this end, we varied the number of exposures and the degree of similarity to other stimuli of visually presented objects. Event-related potentials displayed adaptation effects in all age groups, and were associated with memory specificity. Our findings support the claim that adaptation effects reflect encoding mechanisms that facilitate the formation of stimulus-specific memory representations, and underscore their significance as neural indicators of individual differences in episodic memory across the lifespan (Sommer et al., 2021). The extension of this investigation to children was accomplished in collaboration with Sarah Weigelt (TU Dortmund University). Ongoing analyses use spectral pattern similarity analysis (as described in a recent tutorial by Sommer et al., 2022) to further characterize lifespan age differences in memory representations from encoding to retrieval.
Key References
Research Area 2: Effects of Context on Memory Representations
Successful memory is greatly aided by the spatial and temporal settings of an event, commonly referred to as its context. This dependency of memory on context increases with advancing adult age (Lindenberger & Mayr, 2014). At the same time, and somewhat paradoxically, older adults find it particularly difficult to actively retrieve specific object–context associations. Based on these observations, our studies aim at a better understanding of age differences in the contextualization of memories (i.e., object–context binding) and the precise conditions under which memory performance benefits from context reinstatement (Dissertation Anna Karlsson).
At the neural level, modulations of oscillatory activity (i.e., rhythmic fluctuations in synchronous neural activity) in the alpha and theta frequency band have been related to successful memory encoding and retrieval in young adults, but their role in memory decline in older age is not well understood. With regard to memory formation, we found that theta synchronization and alpha/beta desynchronization remain relevant in older age (Karlsson & Sander, 2022). Alpha/beta desynchronization was less pronounced in older adults than in younger adults, and was accompanied by reduced memory for object–context associations. These observations match similar findings from a study on memory retrieval (Karlsson et al., 2020), where alpha/beta desynchronization was also reduced among older adults and associated with less detailed object memories. Together, these findings suggest reductions in alpha/beta desynchronization with advancing adult age contribute to an impoverished representation of contextual information.
A key candidate for our ability to specifically associate distinct pieces of information during learning is the temporally precise interaction between oscillatory activity in the theta and the gamma frequency band, so-called theta–gamma coupling. We therefore asked whether poorer object–context binding is associated with lower neural coupling precision (Karlsson et al., 2022). We found that gamma power increases are indeed tied to a specific phase angle of the theta rhythm during successful formation of object–context associations in both younger and older adults (see Figure 2). Associative memories were more likely to be retrieved when the gamma power increases were closer to this theta phase angle, suggesting the existence of an optimal theta phase angle for laying down bound memories. Furthermore, for events where only the object was remembered, but not the context, the theta phase at which gamma power increases occurred was shifted away from this optimal theta phase. Finally, adult age differences in associative memory performance were accompanied by a deviation in coupling phase angle further away from the optimal theta phase in older relative to younger adults. In concert, these findings provide evidence for a crucial role of theta–gamma coupling in the construction of a coherent memory trace. They point to decrements in the precision of theta–gamma coupling as a candidate mechanism for the general proposition, put forward by Fergus I. M. Craik and others, that normal cognitive aging compromises the ability to form and retrieve associative memories.
These results are based on a multimodal study that combined EEG and functional and structural MRI with the aim of investigating how context shapes younger and older adults’ memories for objects. In follow-up analyses, we are currently investigating learning-related changes in neural patterns of object-specific representations in the hippocampus as captured by fMRI.
 Illustration of age differences in the distribution of gamma power (y-axis) over theta phase bins (x-axis) for pair memory (left) and item-only memory (right). Shaded areas indicate 95% confidence intervals and phase 0° corresponds to the theta peak (cosine). Relative to younger adults, the coupling phase is shifted in older adults. (b) Illustration of the time (x-axis) and frequency (y-axis) power spectrum within the gamma band (50–75 Hz) for pair memory (left) and item-only memory (right). The black line represents a peak-locked average of the unfiltered EEG signal aligned to the middle peak (time = 0) of the theta rhythm. Coupling accuracy is lower in older adults than in younger adults. (c) The association between the proportion of correct pair responses (y-axis) and individual coupling phase (x-axis; angle in degrees). For illustrative purposes only, absolute phase angles are plotted. Colored lines and dots indicate regression lines for the two age groups and individual data points, respectively. Coupling closer to the peak of the theta phase is associated with better pair memory performance.")
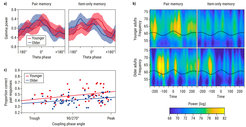
Figure 2. Aging-related decrements in precision of theta–gamma coupling as a candidate mechanism for adult age differences in associative memory. (a) Illustration of age differences in the distribution of gamma power (y-axis) over theta phase bins (x-axis) for pair memory (left) and item-only memory (right). Shaded areas indicate 95% confidence intervals and phase 0° corresponds to the theta peak (cosine). Relative to younger adults, the coupling phase is shifted in older adults. (b) Illustration of the time (x-axis) and frequency (y-axis) power spectrum within the gamma band (50–75 Hz) for pair memory (left) and item-only memory (right). The black line represents a peak-locked average of the unfiltered EEG signal aligned to the middle peak (time = 0) of the theta rhythm. Coupling accuracy is lower in older adults than in younger adults. (c) The association between the proportion of correct pair responses (y-axis) and individual coupling phase (x-axis; angle in degrees). For illustrative purposes only, absolute phase angles are plotted. Colored lines and dots indicate regression lines for the two age groups and individual data points, respectively. Coupling closer to the peak of the theta phase is associated with better pair memory performance.
Image: MPI for Human Development
Adapted from Karlsson et al. (2022)
Original image licensed under CC BY 4.0
Key References